Paradoja de Olbers
Las diatomeas (o Bacillariophyceae) son elegantes, ornamentadas y su desplazamiento resulta un tanto estrambótico. De tamaño microscópico (miden de 10 a 100 micrómetros), las diatomeas son protistas fotosintetizadores, unicelulares, que poseen como característica única una pared celular compuesta de celulosa con una gran concentración de silicio (menos de 56% del peso en seco). En efecto, silicio en forma de óxido (SiO2); como cuarzo traslúcido o ¡vidrio! Todo un suceso evolutivo, que culminó con células contenidas dentro de una auténtica cajita de cristal denominada “frústula”; cajita diáfana, con base y tapa, ambas llamadas hemivalvas (ligeramente cóncavas y unidas por una cintura que permite su separación durante la reproducción). De hecho, la palabra “diatomea” deriva del latín, “di”: “dos”, y “tomo”: “dividido”. Estos microorganismos habitan principalmente cuerpos de agua continentales y, en mayor abundancia, los océanos, donde, en época de elevado contenido de nutrientes, generan auténticos blooms (“florecimientos”, pese a que no son plantas), pudiendo constituir hasta el 70% del fitoplancton. Lo anterior permitiría imaginar al mar brillando en tonos amarillo-pardo (el típico color de diatomeas), así como el gran número de estrellas le permitió a Olbers (y predecesores) imaginar mucha luz y preguntarse por qué las noches son tan oscuras.
Existen registros fósiles de los océanos del Mesozoico que indican la presencia de frústulas de diatomeas silificadas.
El grupo de diatomeas netamente silificadas es joven: surgió hace tan sólo aproximadamente 185 millones de años, durante el Jurásico temprano (Mesozoico), cuando los dinosaurios ya habían colonizado tierras y mares, los helechos predominaban entre la vegetación y los ecosistemas eran de cálidos a ligeramente templados. Los registros fósiles de los océanos del Mesozoico indican la presencia de frústulas de diatomeas con un elevado grado de silificación, tal vez inducido por el alto contenido de sílice disuelto de aquellos mares, pero también de bióxido de carbono (CO2) en la atmósfera mesozoica. De hecho, las diatomeas influyen enormemente en el clima global, en la concentración de CO2 atmosférico y en el ciclo biogeoquímico del silicio. Además, se estima que durante los blooms oceánicos las diatomeas son las principales actrices en la fijación de CO2 vía la fotosíntesis para la producción de materia orgánica y oxígeno, es decir, la llamada “producción primaría neta” (PPN). En ambientes terrestres (suelos y aguas continentales) la PPN debida a diatomeas representa del 20 al 25%, y alcanza hasta el 40% en ambientes marinos.
Fluorescencia, iridiscencia, refracción…
Una de las razones por las cuales las diatomeas nos resultan bellas cuando las vemos al microscopio, es consecuencia de la estrategia evolutiva que significa vivir dentro de frústulas silificadas, de naturaleza “vidriosa”. Y es que vivir así implica que parte de la luz que recibe una diatomea se refracte y parte se refleje, por lo que es un microorganismo con propiedades iridiscentes. Así, las frústulas suponen funciones fisiológicas relacionadas con la fotosíntesis y el uso eficiente de la luz. No sobra decir que, además, la frústula otorga protección a la célula ante factores ambientales adversos como depredación, estrés oxidativo, la acidificación del medio o la presencia de tóxicos, entre otros factores.

Regresando a lo francamente atractivo de estos seres “cristalosos”, es que juegan a su real antojo con la calidad e intensidad de luz que requieren para la fotosíntesis, debido en gran parte a la manera como permiten la entrada y la salida de la luz a través de la frústula. Si miramos bien al microscopio, en la pared celular o frústula de diatomeas podemos observar poros aislados o formando líneas (estrías) que conciertan un rasgo de su bella morfología: la ornamentación. Dado el arreglo ordenado y a escalas meso y nanométrica de poros y estrías se desatan nuevos cánones de “juego” con la luz. En este sentido, nuestros experimentos en torno a las propiedades fotónicas de diatomeas permiten lanzar la hipótesis de que los patrones ornamentales de la frústula (establecidos por el grado de silificación, el tipo y tamaño de los poros) están directamente relacionados con la intensidad y calidad de luz percibidas por las diatomeas. Hemos encontrado en el laboratorio que el tamaño y la forma de los poros, así como el número y el tamaño de cloroplastos y de pigmentos accesorios de la fotosíntesis, cambian en función de la intensidad de luz recibida.
Parte de la elegancia de estos seres microscópicos proviene de que las estrías de las frústulas…
Parte de la elegancia de estos seres microscópicos proviene de que las estrías de las frústulas y los poros que las conforman poseen un arreglo periódico tridimensional: se revela un orden que va desde la microescala (con las estrías) hasta la nanoescala (con los poros). Aún más, este micro y nano orden confiere a las diatomeas propiedades fotoluminiscentes específicas con aplicaciones potenciales para el desarrollo de nuevos materiales y dispositivos electrónicos y ópticos, que permitan almacenar energía o como soporte de catalizadores, sensores o de partículas semiconductoras cual si fueran puntos cuánticos. ¿Diato-tecnología?
La síntesis artificial de dispositivos meso- y nano-estructurados de sílice porosa es costosa, lenta y requiere muchos cuidados y consideraciones. Para hacer más complejo el panorama, esta síntesis artificial resulta contaminante. Ante esto, existen dos soluciones para eludir tales contrariedades: 1. Emplear diatomeas, ó 2. Mimetizar la biogénesis de frústulas. No está demás añadir que los meso- y nano-dispositivos de sílice porosa tienen propiedades fotoluminiscentes de las cuales carece la sílice amorfa no estructurada, por lo que estos dispositivos son esenciales para la construcción de diodos, chips, conductores eléctricos y ópticos, superconductores, celdas fotónicas, supercapacitores, fotocatalizadores, base de pinturas… Es por esto que algunos investigadores están interesados en los procesos biofísicos y bioquímicos que desembocan en la biogénesis de las frústulas (asimilación de Si, metabolismo y regulación a nivel genético de la silificación, por ejemplo). Y es que en el interior de cada célula de diatomeas existen biorreactorcitos denominados vesícula de deposición de sílice (SDV), dentro del cual el ácido silícico, Si(OH)4, se polimeriza. La SDV tiene sus catalizadores proteicos que, además, sirven de “templetes” para la síntesis de cada hemivalva. ¡Figúrese usted que todo ello sucede a presión y temperatura ambiente! Habría que destacar que la ingeniería biomimética está en marcha como una disciplina emergente en un contexto humano deseoso y necesitado de procesos más certeros y sustentables en la producción de materiales útiles, esta vez siguiendo la esencia de las rutas biológicas.
Hologramático
Como en toda interacción, para bailar con la luz se requiere que todos los participantes estén presentes. Sucede que no siempre se tiene suficiente luz… y ello no implica que sea de noche o que la luz esté “apagada”. Tal vez la calidad de la luz no es la adecuada: en cuerpos de agua sabemos bien que la intensidad y el color de la luz varían con la profundidad; en este caso, las diatomeas tendrán que desplazarse a lo largo de la columna de agua que habitan (modulando el contenido de vesículas de gases o grasas intracelulares) hasta donde la interacción pueda establecerse, con lo que se desencadenan las reacciones fotosintéticas dependientes de la luz. La migración de las células no es cualidad única de animales: bacterias, archaeas y protistas (como las diatomeas) igualmente migran, sobre todo si de allegarse a la fuente de energía, como los fotones, se trata. Las diatomeas mantienen una complicidad con la luz (y también con la oscuridad).
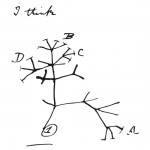
Y de aquí, al baile. Las diatomeas presentan desplazamientos sobre superficies que parecen realizados a “voluntad” y que ocurren de manera extremadamente peculiar. ¡Claro! Evidentemente con una pared tan rígida, de sílice, una diatomea no tendrá un desplazamiento ameboideo o como el tipo “sacacorchos” que se observa en bacterias espirilos. Tampoco será un traslado girando como microorganismos coccoides, mucho menos empleando la propulsión que a espermatozoides les da su flagelo pues las diatomeas silificadas carecen de esta “cola”. Sorpréndase entonces porqué el desplazamiento es casi “gatuno”. Sí, de gato que va rodeando objetos en lugar de seguir una línea recta hasta el “destino final”. Y entonces, ¿Cuál es el destino final? ¡Se trata de trayectorias curvas! En sentido y contrasentido de las manecillas del reloj, alternadamente. Esto es lo que hemos observado cuando diatomeas pennadas del género Nitzschia (fig. 1, ver[Nota 2]) se desplazan en su pista de baile que es el cubreobjetos de un microscopio.

Pero hay más que decir. Durante su periplo la diatomea secreta un filamento de aspecto mocoso, adhesivo, a través de una hendidura conocida como rafe. Ese moquito filamentoso no sólo le permite a la Nitzschia adherirse a la superficie, sino también deslizarse curvadamente. Y siguen los datos: la amplitud de las curvas que sigue la Nitzschia dependen de la forma y la longitud del rafe. Los análisis en torno a este ejemplo de desplazamiento indican que las diatomeas siguen un mecanismo de movimiento estocástico llamado proceso de Poisson, felizmente conocido también como la Ley de los sucesos raros. Y es que este tipo de desplazamiento no se ha observado en otros microorganismos (por ejemplo, bacterias). Para más extravagancia, el desplazamiento no responde a un estímulo externo ni es resultado de una exploración del medio (lo que sí parece suceder en gatos). Los datos de las observaciones y simulaciones realizadas por el grupo de trabajo del coautor le sugieren a la coautora que, más que hacer símiles con un gato, habrá que hacerlo con una araña que teje su “residencia”, en el sentido de que “ahí residirá”, se mantendrá, un rato de su ciclo de vida.
Así entonces, en cada “pasito tun-tun-pasito tun-tun”, curveándose “de aquí para allá” en sentido y contrasentido a las agujas del reloj, la diatomea, que suele jugar con la luz, va dejando un filamento de mucílago que, eventualmente, constituirá la porción extracelular de una biopelícula o biofilms… y hablar de biofilms da para otro artículo con un título tal como “Anarquía y biofilms microbianos”. La pregunta ahora es: ¿Esto es una estrategia diatomeana para un periodo de vida sedentaria? ¿Y qué pasa con la luz a la sazón? ¿Se apaga el foco y se acaba el baile? En eso estamos, mientras tanto, escuchamos el “Pachuco Bailarín” de Pérez Prado. C2
Razones cristalinas para un planeta: CO2 en la atmósfera terrestre
En el Mesozoico la temperatura global era significativamente mayor a la actual, muy probablemente debida a la alta concentración de CO2 (1950 ppmv); la concentración de CO2 disminuyó desde entonces, hasta 260 ppmv en 1850, año fijado como el del fin de la etapa preindustrial. Una elevada proporción de este CO2 se fijó en forma de biomasa. El problema es que a partir de la era industrial, la concentración de CO2 ha incrementado ~1.4 veces; hasta alcanzar 400 ppmv en 2014. No es coincidencia que la actividad metabólica de la creciente comunidad de diatomeas y otros microorganismos fotosintetizadores se relacione con los descensos en el nivel de CO2 de la atmósfera del Mesozoico e inicios del Cenozoico, como no lo es tampoco que el origen de la fotosíntesis desatara la oxigenación de la Tierra, por un lado, y fijación del CO2, por el otro, desde el periodo Precámbrico. Tampoco es producto de la coincidencia, de “mentes adversas al desarrollo industrial” ni de “fuerzas ocultas del neocomunismo ambientalista” (sensu el súper conservador Tea Party) que el CO2 antropogénico afecte la atmósfera, al ser emanado con una aceleración sin precedente en apenas 164 años, vía la quema de combustibles fósiles que contienen las miles de millones de moléculas de CO2 fijadas y metabolizadas por fotosintetizadores. ¡Es mucho CO2 reincorporado a la atmósfera en muy poco tiempo! Lo que explica la preocupación ante el por demás evidente deterioro ambiental global.
Mi forma, mi nombre
Las formas de diatomeas varían de especie a especie, más aún, de género a género… vaya, varían ¡hasta de una Orden a otra! Las hay cocoides, cilíndricas, elípticas, discoides, lanceoladas o lineal lanceoladas¿Cómo clasificarlas? Al puro estilo “escuela-Montessori”: la forma! Las diatomeas con simetría radial pertenecen a la orden de las Centrales; mientras aquellas con simetría bilateral, son las Pennales (del latín penna ‘pluma’, de donde viene la palabra “penacho”). Estas últimas se subdividen en dos subórdenes, Rhapidineae, o rafideas para los cuates, en tanto que tienen rafe (Fig. 1) y Arhaphidineae (sin rafe). ¿Por qué decirlo? es que el rafe, una hendidura que corre a lo largo del eje apical y comunica el interior con el exterior de la célula, juega un papel importante en la movilidad de la célula. Por el rafe, se secreta un moco o mucílago que está implicado en la adhesión y el desplazamientode diatomeas rafídeas, en superficies.

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/
- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

{kind=link}
{kind=link}
{kind=link}