{kind=link}
Bichos, cámara, acción I: Los Fermentadores
Foto de Polina Kovaleva
Cuando células de levadura o bacterias ríen en el jugo de uva
durante la fermentación,
solo ocurren reacciones dionisíacas
Respirar nos es tan común, tan familiar: jalamos aire con su buena dotación de oxígeno molecular (O2) por nuestra nariz, lo llevamos a los pulmones y desde ahí se redistribuye vía arterial para alcanzar todas las células del cuerpo, que las necesitan para, en esencia, “quemar” (oxidar) moléculas y liberar de ellas energía. Sin ésta, ningún sistema biológico subsiste. Así, el O2 es el agente “comburente” (oxidante) que reacciona sobre un “combustible” (moléculas) y, de manera no vigorosa sino controlada (¡no provoca un incendio!), liberará electrones, ¡la fuente de energía! Dicho de otra manera, “respirar” es “quemar alimentos” para liberar la energía contenida en enlaces químicos que establecen los electrones en tales moléculas inorgánicas (p.e., sales de azufre o hierro) u orgánicas (carbohidratos como la glucosa, lípidos, proteínas).
Pero no todos los seres vivos poseen arterias, nariz, ni mucho menos pulmones; solo algunos animales y no en todas las etapas de su ciclo de vida (solo ciertos vertebrados como anfibios y reptiles adultos, además de aves y mamíferos). La inmensa mayoría de seres vivos carecen de alguno de estos componentes del cuerpo (invertebrados, vertebrados cartilaginosos y la mayoría de los peces) o de todos (bacterias, arqueas, protistas, plantas y hongos). Para más, una inmensa diversidad de seres vivos ni siquiera utilizan el oxígeno como agente oxidante (bacterias y arqueas anaerobias, algunos protistas y hongos) … Vaya, ¡no todos los seres vivos respiran! o no obligadamente. ¡Oh!, ¿todo lo que se oxida es porque sucede una respiración? No. “Algo” se puede oxidar abióticamente, pero también bióticamente sin que “eso” sea “respirar” sino ¡fermentar!
Actualmente sabemos que la fermentación es una estrategia metabólica antigua y sencilla de oxidación o descomposición de ciertas moléculas orgánicas (Tabla 1) en ambientes anóxicos o microaerofílicos (con nula o baja concentración de O2, respectivamente), como lo fue la Tierra del Archeano (4000-2500 millones de años); es una oxidación parcial, pues a diferencia de las respiraciones anaerobia y aerobia, no participa algún agente oxidante externo sino que el compuesto orgánico (por ejemplo, la glucosa, C6H12O6) ¡parte se oxida y parte se reduce! (Ec. 1):
1/3C6H12O6 + 2/3C6H12O6 → 2CO2 + 2C2H6O
(Ecuación 1)
Nota interesante: el estudio de la mezcla de enzimas que participa en la fermentación fue motivo del premio Nobel de Química de 1907 para Eduard Buchner, quien la refiriere como un proceso que entrelaza lo vivo y no vivo (así se originó la vida, de hecho) en un jugo de frutas y que puede suceder sin células. ¿Sin células? Sí, pero no sin enzimas, como lo descubrió María Manàsseina [1] antes que Buchner. Los experimentos de Manàsseina “descartaron la teoría fisiológica de la fermentación de Louis Pasteur y confirmaron la hipótesis química propuesta por científicos tan destacados como C. Bernard, J. von Liebig y M. Berthelot” (Kovalzon 2009). Si bien el laureado Buchner se basó y hasta replicó el trabajo de María Manàsseina, opinó que ella solo estaba “subjetivamente convencida de la existencia de una enzima de fermentación” ¡Huy, ciertos hombres de ciencia!
La red de reacciones metabólicas se descompone en módulos y capas (…)
que corresponden a una secuencia histórica de acreciones
Smith y Morowitz, 2016
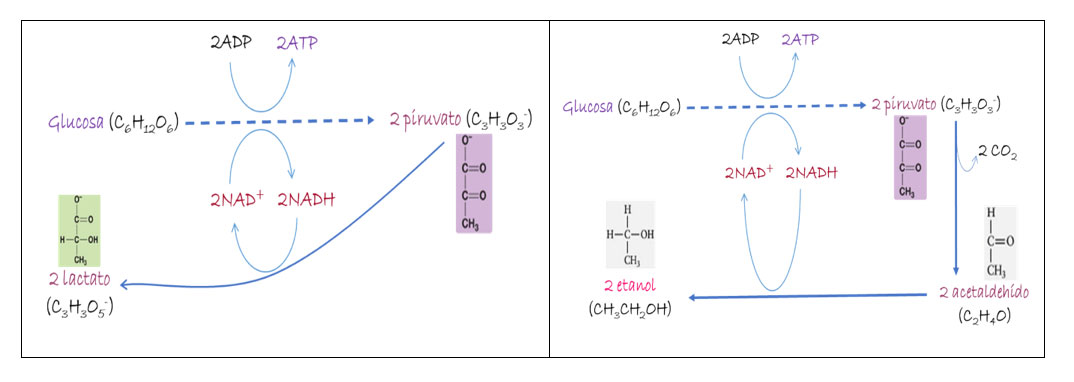
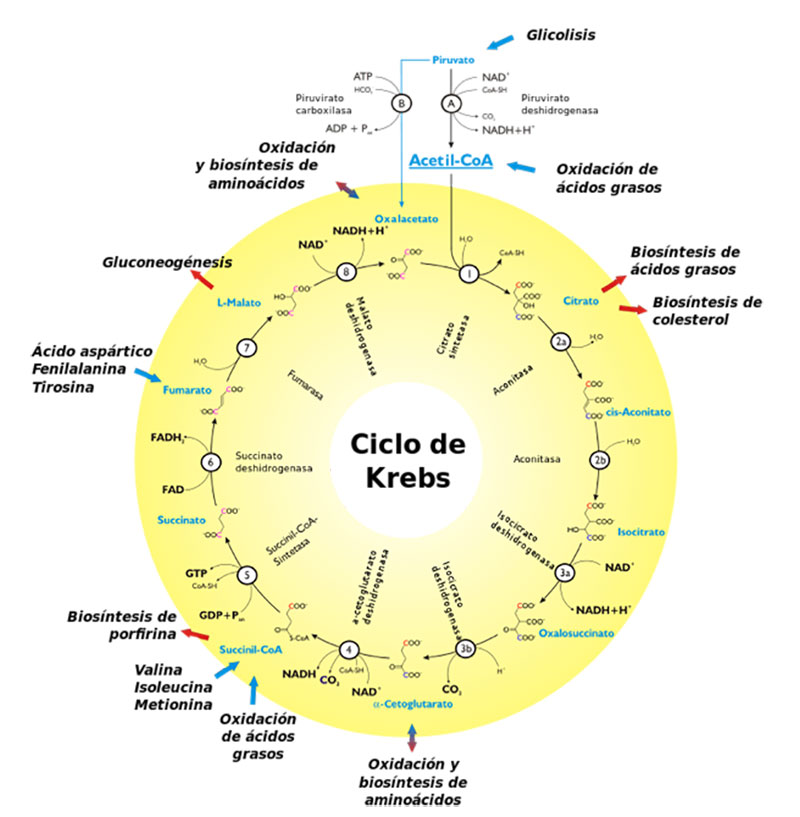
La fermentación es, entonces, una forma efectiva y muy directa de obtener energía y sí, sí se asocia principalmente con la “oxidación” de la glucosa (o glucólisis) que, como vemos en la Ec. 1, resulta ser un proceso metabólico, una fuente de energía que no requiere de agente oxidante alguno. Citando a Deamer (2011), la fermentación glucolítica “es un ejemplo de la forma en que se puede extraer energía de una serie de reacciones químicas vinculadas que podrían ocurrir en la Tierra antes de que el oxígeno estuviera disponible”. En aerobios, sabemos, la oxidación de glucosa presupone el uso del oxígeno, —¿evolucionó este proceso metabólico a partir de un ancestro? Parece ser que sí. La segunda etapa de la glucólisis aerobia genera dos moléculas de piruvato (como los mostrados en la Fig. 1); en ausencia de organelos (p.e., mitocondrias), o de los genes que codifican las enzimas y coenzimas necesarias para llevar a cabo el llamado Ciclo de Krebs (también llamado ciclo del ácido cítrico o ciclo de los ácidos tricarboxílicos; Fig. 2), tales piruvatos no son “respirados” sino fermentados hasta compuestos como ácido láctico o alcohol (Fig. 1). Aquellos con tales “ausencias” son metabolismos tempranos, ancestros de respiradores: la fermentación es un “módulo” al cual se le “adicionó” posteriormente otro, dando origen a la respiración.
Sabemos más: los primeros fermentadores vivieron en ambientes anóxicos, con bajo contenido de agentes oxidantes y, por lo tanto, de bajo potencial oxidante (ca. -0.5 V), así como con cierta dotación de materia orgánica; no obstante, tales condiciones no favorecen la oxidación; gracias a la fermentación estos seres vivos lograron obtener energía, con pocas moléculas de ATP (Fig. 1) al oxidar parcialmente compuestos de carbono orgánico (Ec. 1) utilizando la coenzima NADH para el transporte de electrones e hidrógeno, H (Fig. 1).
Como se mencionó, la fermentación es un tipo metabólico antiguo; hay quienes la consideran, junto con la metanogénesis (p.e., fermentación metanogénica) como el primer tipo metabólico, lo cual es sugestivo si recurrimos al árbol filogenético de la vida de Carl Woese (https://www.revistac2.com/el-arbol-de-la-vida/), en cuyas ramas tempranas de bacterias se ubican como representantes del género Thermotoga, bacterias fermentadoras, aunque también a Aquifex, bacterias quimiolitótrofas (reductora de S). Resulta así que, tanto la fermentación como la quimiolitotrofía son metabolismos antiquísimos, quizá contemporáneos. Y sí, también se les encuentra asociados en ecosistemas naturales o artificiales como la columna de Winogradsky (https://www.revistac2.com/azufre/) en la cual ambos tipos metabólicos conforman la base (Fig. 3). Esta columna parece recrear la historia evolutiva de los metabolismos en la Tierra. Lo notorio es que un fermentador requiere de compuestos orgánicos previamente sintetizados, lo que no permite establecer una teoría de la fermentación como “primer tipo metabólico” (si es que nada más se originó uno) en el origen de la vida. De entrada, diremos que la fermentación sí es de los primeros tipos metabólicos, y a partir de los primeros seres vivos, la vida se diversificó. Veamos el siguiente párrafo.
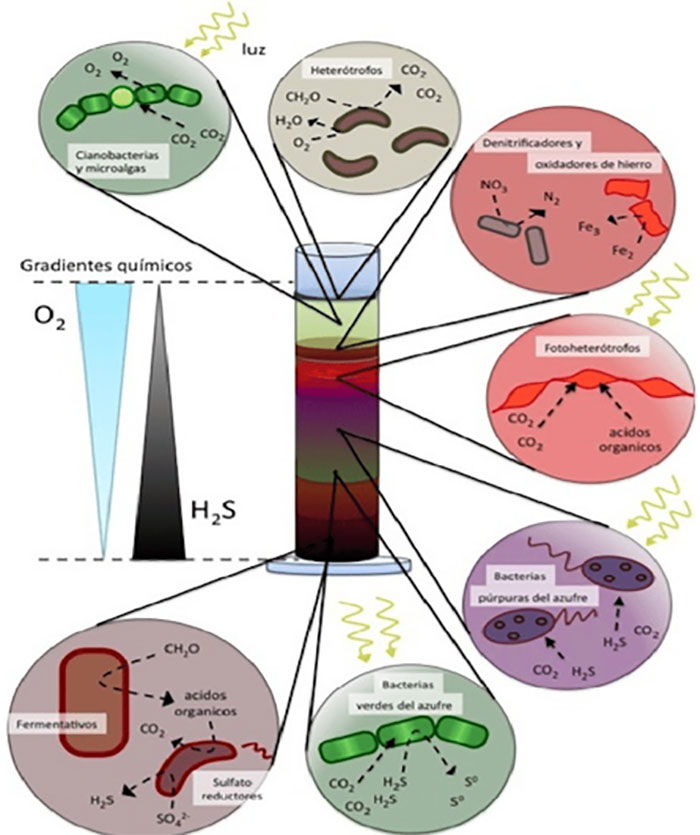
Para la microbióloga Lynn Margulis, el prototipo del primer fermentador es una bacteria tipo Clostridium, dotada de ferredoxinas (https://www.revistac2.com/azufre/) que le permitió tanto fermentar como fijar N2 atmosférico. Aunado a lo anterior, ciertas Clostridium (p.e., Clostridium aceticum) pueden sintetizar compuestos orgánicos como el ácido acético (Ec. 2), gracias también a que poseen proteínas FeS codificadas en sus genomas como las tienen otras bacterias y arqueas (sulfato-reductoras, Fig. 3, y metanógenas) (Sousa et al. 2013). Más aún, alrededor de 355 análisis filogenéticos identifican a clostridios y metanógenos, con estilos de vida modernos similares al del último ancestro común universal (LUCA) o cenancestro (no confundir con “el primer ser vivo” o progenonte), como basales en sus respectivos dominios, habitando en un entorno geoquímicamente activo rico en H2, CO2, S y Fe.
2CO2 + 4H2 → CH3COOH + 2H2O
(Ecuación 2)
Ciertamente, un elevado volumen de la Tierra representa los mayoritariamente abundantes ambientes anóxicos, y donde han poblado fermentadores desde su origen. No obstante, no son las únicas condiciones en las que se pudieron desarrollar los fermentadores, ya que en otras zonas de la Tierra predominaron agentes oxidantes cada vez más fuertes como el citado CO2 y las formas reducidas de Fe y S, estableciendo potenciales redox más elevados, tanto que es viable la presencia del oxígeno molecular, O2; al aumentar y acumularse la concentración del O2 en la atmósfera y en algunas zonas de los cuerpos de agua, estos ambientes se tornaron óxicos; aún ahí, en ambientes óxicos, es posible la presencia de organismos fermentadores facultativos. Un ejemplo de bacteria fermentadora anaerobia es Clostridium y de facultativa, Acetobacter que lleva a cabo la fermentación del alcohol etílico a ácido acético.
Ya sea en ambientes anóxicos u óxicos, la importancia ecológica de la fermentación no es menor desde su origen, al desencadenar la liberación de energía contenida en moléculas y contribuir en los ciclos biogeoquímicos de la materia y el flujo de la energía, así como dar inicio a importantes redes de interdependencia entre organismos (Fig. 3); las bacterias fermentadoras interaccionan entre ellas, con otros microorganismos anaerobios o aerobios e, incluso, con macroorganismos (p.e., fermentadores del tracto digestivo en animales) realizando en conjunto transformaciones químicas para incrementar el valor energético de moléculas mediante las llamadas relaciones sintróficas de degradación para obtener moléculas sencillas, aprovechables tanto como la energía liberada. He aquí la importancia de fermentadores en la biosfera.
Regresando a la fermentación y a Clostridium, este proceso se asocia principalmente con la glucólisis en ambientes anaerobios. Sin embargo, algunas Clostridium llevan a cabo la fermentación de aminoácidos para la síntesis de ATP, en el que uno de los aminoácidos se oxida, generalmente la alanina, y otro se reduce, comúnmente la glicina; este último es uno de los aminoácidos propuestos en la probable reacción de fermentación temprana sugerida por Clarke y Elsden (1980):
glicina + NADH + ADP + Pi-4-acetato + NH4+ + NAD+ + ATP
(Ecuación 3)
Así, otros tipos de fermentación son posibles (Tabla 1) dependiendo de las condiciones del ambiente circundante y, por lo tanto, de los sustratos presentes susceptibles a ser fermentados, y de los miles de microorganismos que lo habiten, resultando diversos productos, algunos de los cuales dieron origen a la llamada “civilización” (que etimológicamente hace referencia a asentamientos humanos), pues posibilitan el uso de compuestos para la elaboración y conservación de alimentos y ¡la realización de festines! (compuestos que ahora llamamos “biotecnologías”), como lo relata el Dr. Moisés Santillán en su escrito “Bondades Microbianas” (https://www.revistac2.com/bondades-microbianas/).
Tabla 1. Diferentes tipos de fermentaciones que se clasifican de acuerdo con el producto generado y el grupo bacteriano involucrado (Tomado de Carbonero,1975) | ||
| Tipo | Principales productos y usos | Grupos Bacterianos involucrados |
| Homoláctica | CH3CHOHCOOH (ácido láctico). Conservación y acidificación de alimentos; en la industria textil y química | Bacterias lácticas como Streptococcus, Pediococcus y Lactobacillus (algunas) |
| Ácida-mixta | CH3CHOHCOOH, CH3COOH (ácido acético), COOHCH2CH2 COOH (ácido succínico) HCOOH (ácido fórmico), CO2+, H2, CH3CH2OH (etanol). Industria del cuero, textil y farmacéutica | Enterobacteriáceas como Escherichia, Salmonella, Shigella, Proteus, Yersinia |
| Butanodiol | Además de los de la ácido-mixta: 2,3- butanodiol. Fabricación de caucho sintético | Aerobacter, Serratia, Aeromonas, Bacillus polymyxa |
| Butírica | CH3CH2CH2COOH (ácido butírico) CH3COOH, CO2, H2. Aditivos alimentarios, saborizantes artificiales, barnices, fármacos y desinfectantes | Anaerobios esporulados (Clostridium) y algunos no esporulados (Butyribacterium) |
| Acetona-butanol | Además de los butíricos: CH3(CH2)2CH2OH (butanol), etanol CH3COCH3 (acetona) CH3CHOH CH3 (isopropanol). Industria alimentaria, cosmética, como desengrasantes o anticongelantes | Algunos anaerobios esporulados (Clostridium) |
| Propiónica | CH3CH2COOH (ácido propiónico) CH3COOH, COOH(CH2)2. Producción de queso suizo, y como conservante | Propionibacteríum, Veillonella |
Es bien sabido que la fermentación juega un papel muy importante para la vida en la Tierra e incluso desde el origen de los primeros tipos de metabolismo, sin embargo, ¡aún hay más! Si tiene curiosidad sobre otras formas de transformar la materia para obtener energía de ella y cómo la Tierra se fue transformando poco a poco hasta lo que es hoy en día (composición atmosférica, seres vivos y sus metabolismos, ecosistemas), ¡no te pierdas los siguientes capítulos!. C2
Bibliografía
- Carbonero P. 1975. Bioquímica de las fermentaciones. Universidad Politécnica de Madrid
- Clarke PH, Elsden SR. 1980. The earliest catabolic pathways. J Mol Evol, 15:333–338
- Deamer D. 2011. First life. University of California Press
- De Gregoris TB, Barroeta B, Esteve-Nuñez A. 2015. La columna bioelectrogénica. Revista Eureka sobre Enseñanza y Divulgación de las Ciencias, Universidad de Cádiz, 12(3), 529-535
- Kovalzon VM. 2009. Some notes on the biography of Maria Manasseina. J History Neurosc, 18:312-319
- Sousa FL, et al. 2013. Early bioenergetic evolution. Phil Trans R Soc. B: Biological Science, 368:20130088
[1] Manàsseina, por ser la primera mujer graduada en la Academia de Medicina de San Petersburgo, recibió un certificado especial autorizándola como “mujer médico”.

- Andrea Almazán Moctezumahttps://www.revistac2.com/autor/andrea-almaz%c3%a1n-moctezuma/


- Elizabeth Romero Martínezhttps://www.revistac2.com/autor/elizabeth-romero-mart%c3%adnez/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/
- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/
- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/
- J. Viridiana García Mezahttps://www.revistac2.com/autor/j-viridiana-garcia-meza/

{kind=link}
{kind=link}
{kind=link}
Leonardo García -
Que hermoso trabajo, siempre es un placer leerlas y más cuando escriben de temas tan interesantes. ¡Felicidades!
Volveré a leer este artículo en compañía de un buen vino para sentirme más en sintonía con la lectura.