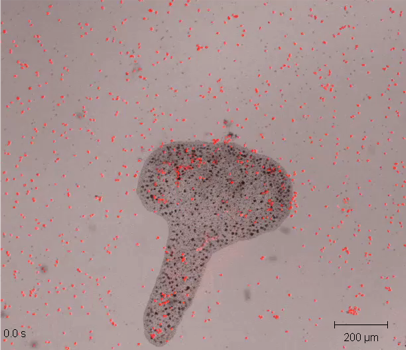
El Trichoplax (Fig.1) es el único ejemplo de animal situado exactamente en el borde de la línea evolutiva entre la vida unicelular y los organismos multicelulares. Es el único miembro del phylum Placozoa. Su genoma es diminuto, pues solamente tiene 12 mil genes, pero identificables en otros animales.
La célula es considerada como la unidad de la vida, y con razón: el sistema ecológico más antiguo y exitoso en este planeta lo forman una variedad de organismos unicelulares, algas, bacterias, protozoarios y hongos (por lo pronto no hablemos de los virus, que muchos no los consideran como seres vivos). Durante más de 3 mil millones de años no hubo otra forma de vida en la Tierra, y persiste hasta nuestros días exitosamente. Células individuales han desarrollado formas muy sofisticadas de regular su metabolismo, alimentarse, moverse activamente y reproducirse.
Entonces surge la pregunta crucial: ¿cuál es la necesidad de la vida multicelular? Creo que la respuesta tal vez está en el excesivo éxito de la vida unicelular. De hecho, si por espacio de unas cuantas horas ninguna bacteria muriera, su población cubriría la superficie entera del planeta con una capa de un metro de espesor. Entonces hay que matarlas. Una solución es el ecosistema mismo, en que una especie se alimente de otra, formando cadenas alimentarias. Pero entre seres unicelulares esto se convierte en una lucha cuerpo a cuerpo, uno se come al otro ¡y nada más!. Otra solución es la proliferación de organismos patógenos, como los virus, que aprovechan las células para reproducirse a miles, matando al hospedero. Ambas estrategias existen en la naturaleza, pero no bastan, se necesita un organismo capaz de matar grandes cantidades de células individuales, es decir, se necesitan súper-depredadores.
Un organismo multicelular es entonces diferente a una simple colonia de individuos.
Entonces podemos ver la aparición de seres multicelulares como una estrategia para mantener a raya a las demás formas de vida. Sin embargo, para que ocurra esto se requieren dos condiciones fundamentales: 1) La división del trabajo, es decir la diferenciación celular, y 2) La organización de todas las decenas de miles de células que forman el organismo por medio de la aparición de sistemas regulatorios y de comunicación que permitan la coordinación de varias tareas, como un sistema digestivo, uno nervioso, uno circulatorio, uno muscular, etc.
Un organismo multicelular es entonces diferente a una simple colonia de individuos. El ejemplo clásico de una colonia muy compleja y que ilustra muy bien el papel del trabajo cooperativo, es el hongo o moho mucilaginoso Dictyostelium discoideum. El moho es un conjunto de amebas individuales que se alimentan de bacterias buscándolas cada una separadamente. Cuando el alimento escasea, se envían señales químicas que las hacen agregarse en un sitio. Al juntarse las amebas (de diez mil a cien mil), forman un pseudo plasmodio, que tiene la apariencia de gusano o babosa y se mueve como si fuera un sólo individuo. Cuando llega a un sitio adecuado, el plasmodio se vuelve redondo y en el centro de su superficie superior aparece una especie de tallo formado por células que secretan celulosa y mueren. Al final, las células que están en la punta del tallo se diferencian en un ‘esporocisto’, del cual surgen esporas, que germinarán cuando las condiciones ambientales sean propicias. Al verlo moverse, uno podría sospechar que el pseudo plasmodio ya es un animal multicelular, pero no es así, pues las células no están fijas en un sitio único en el conjunto, sino que se deslizan continuamente. El movimiento coordinado surge de ondas elásticas al contraerse células individuales por medio de acciones químicas.
El Trichoplax tiene como unas cuarenta mil células, fijas unas a las otras y cada una conserva su diferenciación y su función durante todo el ciclo de vida del animal. Tiene seis tipos de células solamente (Fig2.) y en la mayoría de ellas su función es evidente. Lo raro es que no tiene ningún sistema organizativo, y si lo tiene es muy rudimentario y está oculto. Éstas son (ver referencia (1)):
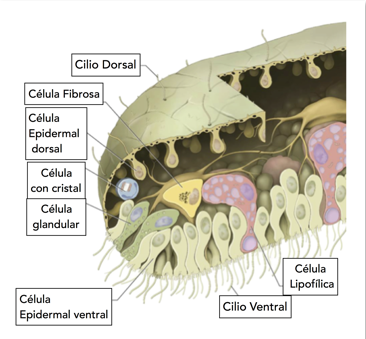
1) Las células lipofílicas, situadas uniformemente en la superficie inferior del animal, secretan enzimas que desbaratan las algas de las que se alimenta, entonces su función es básicamente digestiva. 2) Las células glandulares, poco numerosas y localizadas en la periferia inferior, secretan neuropéptidos: evidentemente son un sistema de señalización química rudimentario. 3) Las células epiteliales dorsales forman una estructura como una membrana que protege y preserva la forma del animal. 4) Las células ventrales son ciliadas (cada una con un pelo, de hecho, Trichoplax significa literalmente ‘plato peludo’) y por supuesto que sirven como un medio de locomoción. El animal vive en los océanos cálidos pero no nada, sino que camina por la superficie inferior o superior del agua. 5) Un poco más misteriosas son las células fibrosas, que se parecen anatómicamente a las neuronas y se conectan con otras células a través del cuerpo del animal. Sin embargo, no existe evidencia de sinapsis ni de conducción eléctrica a través de ellas. Muchos sugieren que es como un esqueleto en ciernes. 6) Aún más misteriosas son las células con cristal, situadas también en la periferia, en las cuales el núcleo central se ha apartado a la orilla y en su lugar está un monocristal de calcita rodeado de mitocondrias. Hay quien sugiere que estos cristales son un sensor de gravedad (como un lastre) que le dicen al bicho para dónde es abajo. Pero sabemos que la calcita es un material birrefringente (con dos índices de refracción) que sugiere que su función es óptica, sugerencia que se reafirma al constatar las superficies convexas paralelas que tienen los cristales (forma parecida a una lente). Sin embargo, el tamaño de los cristales es muy pequeño para que pueda funcionar con luz visible, pero en el ultravioleta la distancia focal se estima como en medio milímetro: ¡el diámetro aproximado del animal!, ¿Podría ser esto un ojo primitivo?
¿Podría ser esto un ojo primitivo?
Ha surgido recientemente un interés renovado por estudiar este bicho, tanto en la biología tradicional, como en la biología molecular, la bioquímica y la genética, pero no existen estudios que se enfoquen en explicar los mecanismos por los cuales este animal pudo resolver el cambio brutal de escala que representa el coordinar el funcionamiento de miles de células, que seguramente son muy básicos y susceptibles de ser modelados con ecuaciones simples, lo que lo hace un sistema fascinante para un físico.
Aquí me gustaría dar un ejemplo de cómo un físico ataca un problema biológico. Empecemos por lo más simple, explicar cómo se mueve el animal. Las observaciones indican que los cilios se mueven periódicamente en lapsos alternados de activación y reposo (ver referencia (2)). Pero medidas detalladas muestran que el movimiento de cada cilio es al azar, más aún, en ausencia de comida el desplazamiento cuadrático medio del centro de masa del animal crece linealmente en el tiempo, que es la característica de un caminante al azar. Entonces la pregunta es ¿cómo es posible que un conjunto de remos que se mueven en direcciones arbitrarias produzcan un movimiento coherente de todo el animal y dirigido para buscar comida?
El animal no nada en el agua, sino que camina, entonces todos los modelos de locomoción basados en la hidrodinámica y desarrollados para amibas, paramecios, espermatozoides, bacterias, e incluso para el reptar del plasmodio de Dictyostelium, son inadecuados en este caso.
La fuerza motora viene exclusivamente del movimiento al azar de los cilios, que al adherirse a la superficie y contraerse producen un pequeño desplazamiento de cada célula en una dirección arbitraria (ver Fig. 3). Si las células estuviesen libres y no interactuaran entre sí, el diámetro del animal crecería como la raíz cuadrada del tiempo (un conjunto de caminantes al azar), pero las interacciones mecánicas entre las células no permiten que cada desplazamiento sea independiente. Existen interacciones de corto alcance: las células vecinas a cada una estorban el libre movimiento individual, y lo que esto produce es una especie de conducta colectiva de enjambre (swarm behaviour) en la que cada individuo trata de adquirir la velocidad promediada de sus vecinos.
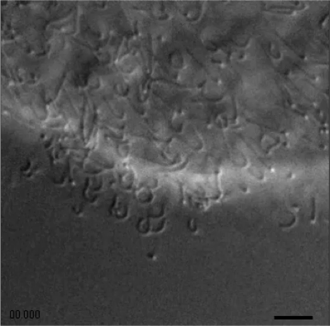
Esto provocaría la deformación excesiva del animal, ya que grupos pequeños de células se moverían en direcciones divergentes, pero entonces las células epiteliales dorsales entran en acción, ya que actúan como una membrana elástica que impide deformaciones arbitrarias del conjunto. También podría proponerse que una de las funciones de las células fibrosas es la de comunicar elásticamente a células más lejanas, y entonces la interacción tiene mayor alcance que simplemente por contacto. Un modelo dinámico con estos ingredientes, desarrollado por nosotros, nos ha permitido reproducir las deformaciones que se observan cuando el animal se mueve.
Sin embargo, el bicho busca la comida.
Sin embargo, el bicho busca la comida. Entonces, proponemos que las células responden a señales químicas que secretan las algas de las que se alimentan. Las algas ‘huelen’, es decir, difunden alguna substancia química que es detectada por las células del animal. Esto no es muy descabellado, muchas células conocidas presentan quimiotaxia, que quiere decir que responden a gradientes de substancias químicas. Si agregamos un término quimiotáctico a las ecuaciones de movimiento del modelo, de forma que se produzca movimiento preferentemente en la dirección de la fuente química, podemos reproducir el movimiento observado en los animales reales. Es casi mágico ver que la conducta del animal parece la de un ser inteligente buscando activamente la comida, y no es más que el resultado de unas pocas reglas que obedecen la física más elemental.
Vemos claramente lo que se ha dado por llamar la aparición de una ‘propiedad emergente’ en un sistema complejo, en este caso una especie de inteligencia artificial. La lección que debemos recordar es que aún los procesos más complicados en los seres vivos tienen que emanar de interacciones simples puramente descritas por la física, y que la investigación en biología se ve enormemente reforzada si no se excluye el modelaje matemático basado en conceptos físicos.
Es por esto que considero al Trichoplax como una cumbre del devenir evolutivo y enfatizo que los procesos evolutivos tienen que tomar en cuenta no sólo las mutaciones genéticas, la adaptabilidad y la selección natural, sino las posibilidades de cambio de los estados estables de un sistema complejo.
Como puede constatarse, la forma en que el animal se mueve es altamente ineficiente, lo que sucede frecuentemente en la evolución. Las soluciones a un problema evolutivo no son nunca perfectas ni únicas, pero perfectibles. Basta con que el mecanismo funcione y permita la sobrevivencia. Vemos por ejemplo que las esponjas (Parazoa), muy cercanas a los Placozoa, no se mueven para buscar comida, sino que aprovechan que el agua la transporta y entonces desarrollan poros y canales por los que la comida circula. Y ambas soluciones para alimentar a un animal multicelular en forma simple han funcionado por millones de años.
Como puede sospecharse, una vez enganchados en este interesante y atrayente problema, es difícil dejarlo, pues siempre hay algo nuevo por estudiar, explicar o descubrir. Esto lo demuestran Carolyn Smith y Thomas Reese, quienes han estudiado el mismo bicho (clones del mismo ‘strain’) desde 1979, y a quienes estoy agradecido por introducirme al fascinante mundo de los Placozoa. Tambien agradezco a Tzipe Govezensky por su colaboración en el modelaje y su asistencia en el tratamiento de los datos experimentales. C2
___________________________________________________
Referencias:
(1) Smith, Carolyn L.; Varoqueaux, Frédérique; Kittelmann, Maike; Azzam, Rita N.; Cooper, Benjamin; Winters, Christine A.; Eitel, Michael; Fasshauer, Dirk; Reese, Thomas S. (2014). “Novel Cell Types, Neurosecretory Cells, and Body Plan of the Early-Diverging Metazoan Trichoplax adhaerens’’. Current Biology. 24 (14): 1565–1572.
(2) Ueda, T., S. Koya, and Y. K. Maruyama. (1999). Dynamic patterns in the locomotion and feeding behaviors by the placozoan Trichoplax adhaerence. Biosystems 54:65-70.

- Rafael A. Barriohttps://www.revistac2.com/autor/rafael-barrio/
- Rafael A. Barriohttps://www.revistac2.com/autor/rafael-barrio/
- Rafael A. Barriohttps://www.revistac2.com/autor/rafael-barrio/

- Rafael A. Barriohttps://www.revistac2.com/autor/rafael-barrio/

{kind=link}
{kind=link}
{kind=link}
Pedro -
12mil genes no son pocos. Se puede comparar con la mosca de la fruta que tiene 14mil.